球孢白僵菌诱导番茄增强对烟粉虱的抗性
植物受害虫、病原体侵袭时会激活局部及系统防御反应,有益微生物如根际微生物和虫生真菌可诱导该防御。球孢白僵菌(Beauveria bassiana)作为常用生物防治虫生真菌,其侵染昆虫的致病机制已有相关报道,但以往研究聚焦其对害虫的致病性,却忽视植物 - 虫生微生物 - 害虫的互作关系。微生物不仅与昆虫存在关联,还能深刻影响植物和昆虫互作过程。研究已证实,虫生真菌蜡蚧轮枝菌(Verticillium lecanii)和金龟子绿僵菌(Metarhizium anisopliae)可通过改变植物代谢成分来刺激植物,诱导昆虫产生趋性或忌避反应。
烟粉虱作为一种广食性植食性昆虫,通过取食韧皮部汁液危害作物,并可能通过其唾液蛋白铁蛋白 BtFer1 抑制植物的诱导防御反应,包括降低茉莉酸(JA)介导的防御信号通路的诱导水平。因此,增强植物防御能力并消除害虫干扰至关重要。植物次生代谢物是植物自身合成的、不影响正常生长发育的化合物,可作为植物防御功能的直接指标,酚类物质是分布最广泛、最常见的成分之一。当植物应对昆虫取食胁迫时,苯丙烷代谢通路被激活,介导酚类物质的合成,进而使活性氧含量升高以抵御害虫。茉莉酸(JA)是植物防御反应的核心调控因子,可响应胁迫并激活防御基因表达、提升防御酶活性。已有研究显示球孢白僵菌内生菌株及同类虫生真菌可增强植物抗虫性,但机制尚不完善。因此,研究人员旨在验证球孢白僵菌可刺激植物对烟粉虱胁迫产生更快、更强的响应假说。

研究方法与结果
基因表达分析
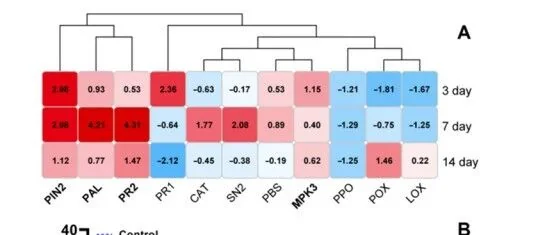
研究采用实时荧光定量 PCR 技术检测球孢白僵菌诱导的植物主要防御通路。通过植物抗氧化试验评估植物的防御能力。实验结果显示:
-
• 处理 7 天后 :番茄防御基因表达显著上调,且效果优于 3 天和 14 天处理组。此时上调的基因数量最多、上调倍数最高,番茄防御水平达到峰值。 -
• 持续上调基因 :PIN2、PR2、PAL 和 MPK3 四个基因在三个时间点均持续上调,其中苯丙氨酸脱氨酶基因(PAL)表达量极高,在 7 天处理时达到峰值,上调 18.5 倍 。 -
• 烟粉虱取食诱导 :与真菌接种处理相比,烟粉虱取食诱导的基因表达水平较低,仅少数防御基因(包括 PAL、POX、PIN2、LOX、CAT 和 PR1)出现上调,基因平均表达水平为对照的 2 倍 。 -
• 联合处理 :真菌接种与昆虫取食联合处理的植株,其基因表达谱呈现出显著差异,其中有 9 个基因上调,且上调幅度均超过 10 倍 。该处理下的防御响应基因主要集中于初级和次级代谢通路,其中 4 个上调基因(PPO、PIN2、PR2 和 PR1)被证实参与酚类化合物的合成。
代谢物分析
通过对四种处理组的 GC-MS 检测结果监测分析发现,结果存在显著差异。代谢物数据基于 KEGG 通路数据库进行比对分析,发现植物始终以营养物质合成代谢为主要代谢途径。具体结果如下:
-
• 球孢白僵菌单独处理 :植物次生代谢物含量急剧增加。经化合物数据库比对,这些代谢物富集于防御通路和激素网络调控通路,具体包括:苯丙氨酸代谢、亚油酸代谢、α - 亚麻酸代谢。 -
• 烟粉虱单独取食处理 :诱导的代谢通路包括:苯丙氨酸代谢、α - 亚麻酸代谢、糖酵解 / 糖异生、乙醛酸和二羧酸代谢。 -
• 真菌与昆虫复合处理 :显著激活了更多防御相关通路,其中由 PAL 基因介导的苯丙烷代谢通路表现得最为丰富和突出。此外,参与酚类等防御物质合成的关键前体物质 — 苯丙氨酸,其生物合成通路也被显著富集。同时,检测到参与植物防御调控的植物激素茉莉酸及其合成前体物质 α - 亚麻酸和 L - 异亮氨酸。经归一化峰面积计算,球孢白僵菌接种处理后,这些物质的含量略有上升。经真菌与昆虫复合处理,其峰面积迅速升至 6.6×10⁴ ,为处理前的 2.4 倍 (p < 0.001)。
抗氧化能力分析
植物的抗氧化能力强弱与酚类代谢物水平直接相关。实验结果显示:
-
• 球孢白僵菌接种 :可轻微提高番茄的抗氧化能力。 -
• 烟粉虱取食植株 :抗氧化物质含量仅升至 0.47 ± 0.0115 mM 。 -
• 茉莉酸甲酯(MeJA)处理组 :抗氧化物质浓度显著升高,为 0.81 ± 0.0036 mM 。 -
• 真菌与昆虫联合处理组 :抗氧化物质浓度显著升高,为 0.79 ± 0.0075 mM 。该处理组叶片的染色试验也显示出最深的颜色;对照组的抗氧化物质浓度仅为 0.47 ± 0.0115 mM ,叶片几乎无明显显色。
结论
综上所述,球孢白僵菌可刺激番茄合成防御物质,引发与烟粉虱取食诱导类似的反应。然而,要充分理解虫生真菌与植物之间的相互作用机制,以及它们在调控害虫与植物互作中的潜力,仍需进一步研究。后续研究可基于几丁质和 β - 葡聚糖等病原相关分子模式与植物模式识别受体之间的相互作用分析,进一步阐明植物对虫生真菌的防御识别机制。尽管该研究在防御识别模式方面存在局限性,但该方法显示出的应用潜力仍然相当可观。

图 1 番茄防御基因的表达模式

图 2 差异基因的 KEGG 富集 通路